
 This report is an edited and condensed version of the report produced by Rachel Cook as part of her final
year project on the Wood Ants of Pound Wood. The project began in the summer of 1997 and was completed in
Spring 1998. It omits much of the detailed survey work, soil analysis and references, but presents the
results that would be of interest to anyone interested in the study of the fauna of Pound Wood.
This report is an edited and condensed version of the report produced by Rachel Cook as part of her final
year project on the Wood Ants of Pound Wood. The project began in the summer of 1997 and was completed in
Spring 1998. It omits much of the detailed survey work, soil analysis and references, but presents the
results that would be of interest to anyone interested in the study of the fauna of Pound Wood.ABSTRACT
The distribution of the wood ant nests was surveyed within 22 hectares of ancient woodland. The relationship between nest density and ecological features was examined to determine the criteria for nest site selection within the wood. The composition of soil samples was analysed, and moisture content calculated to see if ants preferred well drained soils on which to construct their nests. Light intensity was measured to determine the influence of sunlight on nest site selection. The spatial pattern of nests within the wood was analysed to see if the position of neighbouring nests influenced nest site location. In addition, the relationship between nests and species of trees which were foraged for food, was examined to determine their influence on nest site selection. In an ancient woodland the availability of sunlight was shown to be crucial to nest site selection. Nests of Formica rufa were constructed within coppiced areas, along paths or on the edge of woodland. All nests were found on well drained soils, and within areas with access to oak trees which were foraged for invertebrates. The presence of tree stumps or other structures on which nests were constructed, was also found to influence the distribution. In the area where nest density was highest, the position of neighbouring nests also influenced nest site selection by Formica rufa.
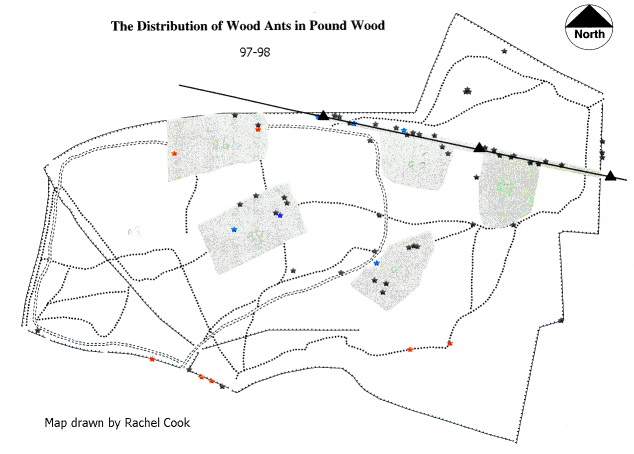
The Distribution of nests of Formica rufa in Pound Wood 1997–1998.
3. RESULTS
3.1 DENSITY AND DISTRIBUTION OF NESTS
At the time of the first survey before the over-wintering period, there were 53 active nests within Pound Wood, and an additional 6 nests which were not occupied. After the second survey during April, the total number of nests rose to 66. There were 12 new nests, 5 of which had been previously unoccupied. Therefore, in April there were 65 occupied nests and one empty nest within the 22 hectares of Pound Wood. The density of nests in Pound Wood during spring was 2.95 occupied nests per hectare. The distribution of Formica rufa nests between August 1997 and April 1998 is shown in the map above. Nests tended to be found in coppiced areas, along the edge of the woodland or along paths next to a coppiced area. The figures for these are given in the table below:
|
COPPICE
|
|||||||||
|---|---|---|---|---|---|---|---|---|---|
| Paths | Chippings | Hedge | Other | WOODS | HEDGE | PATH | TOTAL | ||
| Small (<1m) | 1 | 0 | 0 | 0 | 1 | 4 | 4 | 15 | (23%) |
| Medium (1–2m) | 6 | 3 | 1 | 15 | 4 | 6 | 3 | 38 | (58%) |
| Large (>2m) | 0 | 4 | 0 | 7 | 0 | 0 | 1 | 12 | (19%) |
| TOTAL | 7 | 7 | 1 | 27 | 5 | 10 | 8 | 65 | |
| (11%) | (11%) | (1%) | (42%) | (8%) | (15%) | (12%) | |||
In Pound Wood, 65% of the occupied nests were found within coppiced areas which constitute approximately 1.87 hectares of the total area of the wood (Smart, Essex Wildlife Trust). This gave a density of 22.45 nests per hectare. This is considered a particularly high number, compared to previous studies (Welch, 1978); the lowest densities of nests or where no nests were found, were in wooded areas such as secondary woodland and ‘Non Intervention Areas’.
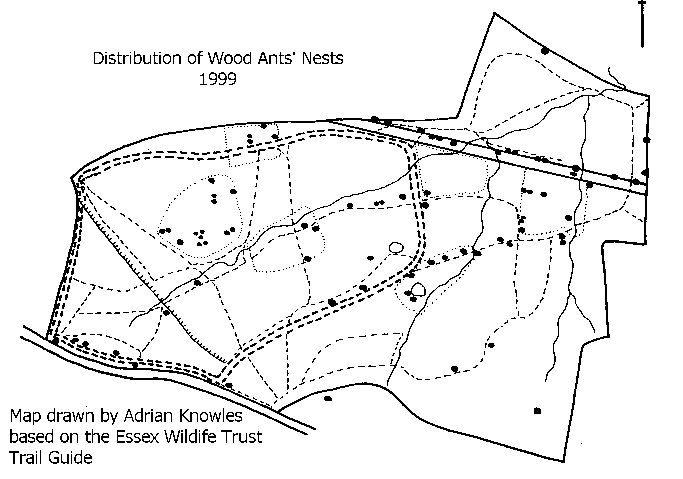
See also the nest distribution established by Adrian Knowles during a survey in 1999.
{kind=link}
3.2 NEST SHAPE AND SIZE
The nests of Formica rufa within Pound Wood showed a variation in shape in response to their location. Nests that were found along a hedge or woodbank were often irregular collections of twigs, usually constructed down a sloping wall. Nests constructed within a wooded area, and receiving more shade such as in the northern section, were taller (to utilise the available radiation). Nests that were positioned along paths were distinctly more flattened (direct sunlight hitting them), although the latter could have been as a result of intervention from passers by. The majority of nests within Pound Wood were of medium size. However, the biggest nests were found within the coppiced areas, and were constructed from wood chippings left in these areas.
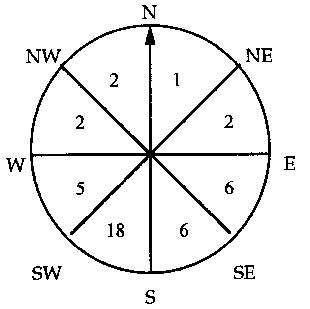 3.3 EXPOSURE
3.3 EXPOSURE
Nests were recorded facing all octants, but the direction of greatest slope was facing SSW in the majority of cases (see Figure on right), increasing the amount of solar energy collected by the nest. Nests always faced the direction of maximum sunlight, unless obstructed by brambles or grass. For example, in the Little Valley coppiced area which was composed of scrub (being cut in 1994–95), there were four nests in 1997. Two nests were found along the northern edge facing south into the clearing in, and two additional nests were found along the eastern border facing west into the clearing, and the sun. In the area under the pylons, nests are lined up along the northern side of the sloped ground, facing South.
3.4 NEST SITE PREFERENCE
Of the 65 occupied nests studied, 51 (78%) were based on rotting tree stumps or fallen branches. The ants used the stumps to base their nests on, the rotting wood being easier to excavate than the compact soil. The wood ants were also seen to utilise the mounds of wood chippings that had been left in the coppiced areas — seven nests (11%) had been built in these. Not surprisingly, all the nests within the coppiced areas were built on tree stumps. Four nests (6%) were built against hedgerows that had been constructed out of layered branches by the Essex Wildlife Trust to separate the paths from wood and coppiced areas. For the three remaining occupied nests it was not possible to determine on what they had been constructed.
3.5 NEST ACTIVITY
Within Pound Wood Formica rufa was active until the beginning of December 1997, with activity above ground resuming on 2nd February 1998 (Smart, Essex Wildlife Trust).
In Topley’s before the coppicing was carried out in February 1998 (which involved felling several large Oaks) the ant nests in the area were active. However, after coppicing three nests (two along paths, and one along the western edge) appeared abandoned — no ants could be seen on the nests. This was the case for three weeks in which the wood was visited weekly, with activity resumed after this period. It was thought that perhaps this event was caused by a disruption to the food supply of the nests caused by the coppicing.
During March and April a tremendous amount of activity occurred in the way of new nest construction. All but one of the nests that were abandoned during the first survey were re-established as active nests during spring. During March several transient nests were occupied on the hedge facing Bramble road. They were later abandoned, probably in response to the extensive growth of foliage in the area.
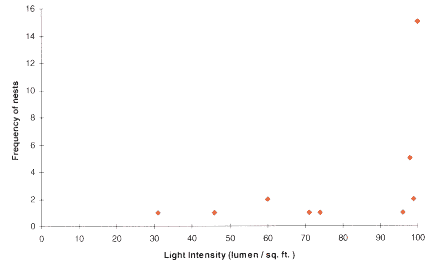
3.6 LIGHT INTENSITY
It can be seen from the figure above, that the majority of nests were located in the areas of light intensity approaching 100 lumen per sq.ft (which was the highest reading on the photometer). However, nests were found under dense canopy, suggesting that Formica rufa can tolerate a certain degree of shade. It can been seen from the graph that there was a minimum light requirement of 30 lumen per sq.ft.
3.8 SPATIAL NESTING PATTERNS
From observations during the mapping surveys it became apparent that there were four types of spatial nesting patterns within Pound Wood. The first was random, where nests were located independently of one another, which was characteristic of low density areas. The second pattern was clumped, where nests were found in clusters such as in northern section (which were satellite nests). The third was where nests were placed solitarily, and the fourth pattern was found in the area under the electricity pylons, where nests appeared evenly spaced apart.
The results of the Clark and Evans (1954) nearest neighbour statistical test on the nesting pattern in four coppiced areas are given in the table below.
|
Area
|
R | Probability |
|---|---|---|
|
Area under the pylons
|
1.162 | <0.05 |
|
Little Valley
|
0.873 | >0.05 |
|
Rowley's
|
1.0508 | >0.05 |
|
Digby's Dell
|
0.674 | <0.05 |
These results show that within the coppiced areas there is a variation in the spatial pattern of nests. The distribution of nests in the area under the pylons is significantly different from a random distribution, and statistically proving that there is an over-dispersed nest pattern in this area. The observed distribution of nests within Little Valley and Rowley’s is not significant at the 5% level, so the null hypothesis is accepted that in these areas there is a random distribution. The pattern of nests within Digby's Dell is aggregated.
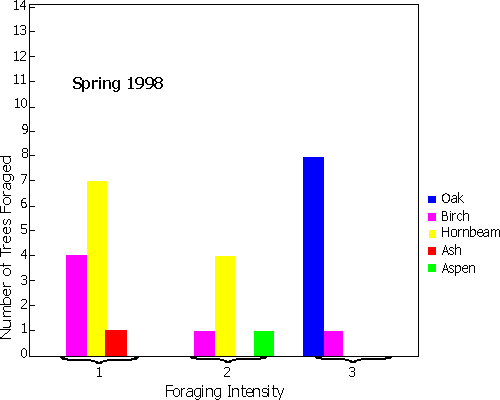 The seasonal
variation in utilisation of trees for foraging by Formica rufa in Pound
Wood. Foraging intensity was measured on a 3 point scale: 1) few ants; 2) faint trail; 3) distinct trail
or many ants. The alternative charts represent
Spring and
Autumn. (Click one of the links above to see the image.)
The seasonal
variation in utilisation of trees for foraging by Formica rufa in Pound
Wood. Foraging intensity was measured on a 3 point scale: 1) few ants; 2) faint trail; 3) distinct trail
or many ants. The alternative charts represent
Spring and
Autumn. (Click one of the links above to see the image.)
The Oak trees (Quercus spp.) in Pound Wood were found to be an important source of invertebrate food for the wood ants. There was a high dependence on these trees all year around (Figure 3.4). The trees were utilised in preference to other trees even in areas where there was a low density of Oaks, such as in the northern section of the woods. Very rarely were the nests of Formica rufa constructed at the base of an Oak tree, but were usually a short distance away with distinct foraging trails leading to several Oak trees (see below). Trails were seen measuring up to 54 m in length.
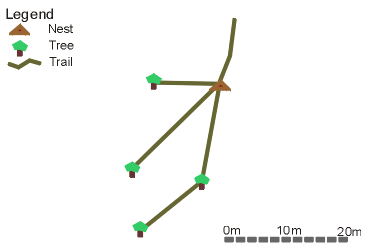
The trails emanating from one
nest.
During autumn no ants were seen foraging on the Hornbeam (Carpinus betulus). However, in spring ants started to appear on them, possibly due to the presence of invertebrates. No ants were seen on the Sweet chestnut trees (Castanea sativa) that are commonly found throughout the wood. In addition, aphids were seen to be tended on Aspen (Populus tremula) under the electricity pylons.
5. DISCUSSION
The general distribution of Formica rufa nests within Pound Wood is similar to previous studies based on the Red Wood Ant in South England. Most have shown a low overall density of nests in comparison to overall area of wood, with nests typically occupying coppiced compartments constituting a small proportion of the woods (Welch, 1978).
Within Pound Wood the nests of Formica rufa were predominant in areas of highest light intensity, namely the coppiced plots, paths or along the edge of woodland. To maximise the radiation received, nests in Pound Wood were always orientated towards the sun which was South-facing in the majority of cases. Laine and Niemela (1989) found that nests occurred on North-facing slopes, providing that the local relief offered favourable southerly exposures; Yarrow (1955) states that nest shape is a function of the habitat and not the ant. In Pound Wood the workers of Formica rufa were shown to craft their nests to response to the incidence of infra-red rays. The fact that the largest nests were found within the coppiced areas reflects the importance of light for the success of nests. Brian and Brian (1951) found that both the size of the colony and the size of workers grew in response to an increase in radiation. It can be seen that the results of the study in Pound Wood support the hypothesis (Adams, 1993) that the availability of sunlight greatly affects the distribution of Formica rufa.
Coppiced areas were favoured not only because of increased radiation received by the colony, but also because the abundance of invertebrates (which provide protein for the development of the brood) increased with the growth of herbs in areas where sunlight was plentiful. Welch (1978) found that young mixed coppice provided greater numbers of both species and individuals as prey for Formica rufa. Another factor that may have influenced the distribution of Formica rufa within Pound Wood, was the coppice cycle and age of vegetation within the coppiced panels. Welch (1978) also found that during the first two years after coppicing there was a rapid increase in nest density in response to the growth of herbaceous flora and availability of prey. This was followed by a steady decline in nest density as the canopy grew over. A similar pattern of colonisation was seen in Pound Wood. In Rowley’s (which was coppiced between 1996–97) two new nests were seen in March 1998, one was a satellite nest formed from a nest on the other side of the path. In the other coppiced areas, only the previously unoccupied nests were re-colonised in spring. Little Valley, was coppiced five years ago, and in this area the distribution of nests was limited to paths and the edge of the coppice because the interior was covered by Brambles which commonly dominate the fifth year after coppicing (Welch, 1969). Brian and Brian (1951) found that wood ants were rarely seen living beneath Brambles because of the dense shade that they cast. In addition, there were no new nests within the coppiced panels of under one year, where the main sources of food had been removed.
Oak trees within Pound Wood were found to be a fundamental source of food for the wood ants. In an analysis on the species of insects associated with British trees, Kennedy and Southwood (1984) found that Oak trees harboured 423 species of insects, which was the highest number recorded. In Pound Wood Formica rufa does not construct nests in close proximity to these trees; however nests were always found in areas where there was access to Oaks via foraging trails. The description of foraging activity given by Skinner (1980a), who states that ‘active ant roads ran from each nest to a nearby Oak tree and then on to other Oak trees, and could attain a length of 53 metres or more’, is similar to the activity seen in Pound Wood. The northern section of the wood consists mainly of Hombeam which was found to cast deep shade, and was poor in ground vegetation. However, in this area there were four thriving nests. Oaks were found at a very low density, but were foraged by ants all year which may have been an important factor in the success of the nests. The findings of this study do not support the hypothesis (Adams, 1993) that Oak trees directly affect the distribution of Formica rufa nests. However, the success of Formica rufa within Pound Wood can be attributed to the presence of these trees.
Mahdi and Whittaker (1993) found that foraging by Formica rufa on Birch (Betula pendula) was lowest between May and June when most of the foraging activity focused on Oak trees. The results of the study in Pound Wood also showed that different species of tree were foraged by Formica rufa at different times of the year. However, in contrast to the study by Mahdi and Whittaker (1993), Hombeam and Birch were foraged early in the year, and Oak all year. This may have been related to varying densities of invertebrates found on these trees throughout the year. Hill and Roberts (1990) found that Birch and Hombeam supported up to 12 times higher densities and biomass of invertebrates than Sweet chestnut, which may explain why no ants were seen foraging the Chestnut trees.
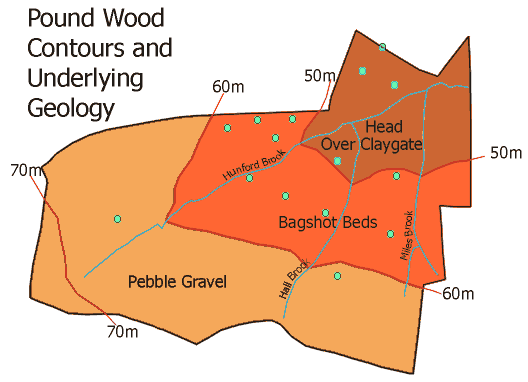
Within Pound Wood nests of Formica rufa were found on soils containing varying amounts of sand, silt and clay (the green spots on the map above). However, the soil samples were all categorised as Loams, implying that they were of similar texture, so all areas should have been from the point of soil composition, equally favourable to Formica rufa. The absence of Formica rufa from certain areas may not have been attributed to the soil in those areas. The lack of nests on the plateau gravel may have been as a direct result of another factor such as the absence of suitably lit areas in the Non-Intervention Areas of the wood where the plateau gravel is found. In the soil samples taken from the wood there was good drainage of water which is a prerequisite for nest excavation by Formica rufa. Therefore, the results failed to support the hypothesis that the composition of soil is an important factor in nest site selection of wood ants (Adams, 1993). Instead the results of the study support the hypothesis that soil type is not unduly important as long as drainage is good (Yarrow, 1955).
Levings and Traniello (1981) found in their studies on spatial patterns in ant populations, that when there were low densities of nests the distribution of nests reflected the best foraging or nest sites, and nests were dispersed randomly. The results of the nearest neighbour statistical test (Clark and Evans, 1954) showed that in both Little Valley and Rowley’s coppiced panels there were random distributions of nests. In Dibgy’s Dell the pattern of nests was more aggregated, possibly because the area had been colonised by the establishment of satellite nests. Nest density was highest in the area under the pylons where nests were constructed amongst an established community of herbs and grasses with access to surrounding wood, and received a great deal of sun. The nests in this area showed a more evenly spaced or more over-dispersed distribution than a random one. Wilson (1971) states that over-dispersed patterns of nesting are common among ant colonies of the same species when intraspecific competition occurs for resources. Ryti (1990) found that nest spacing of Formica allipetens became increasingly regular on sites where the density was over 20 nests per hectare. This corresponds with the findings in the area under the pylons where the density was 28 nests per hectare. The results support the hypothesis (Bemstein and Gobbel, 1979; Levings and Traniello, 1981) that in an environment where physical conditions for brood development are uniformly available (such as the area under the pylons), the position of nests will depend on the location and foraging areas of neighbouring nests, and within such an area nests will be more regularly spaced than randomly. The density of nests within the area under the pylons would be expected to increase to the point of carrying capacity, and after that point other areas would be colonised if available.
In Pound Wood the hedgerows laid by Essex Wildlife Trust, and tree stumps on which nests were constructed, were found to influence the nest site selection of Formica rufa. Adams (1993) found that out of 300 nests, all were constructed upon rotting tree stumps.
The nests of Formica rufa within Pound Wood showed a great deal of movement during the study period. In spring swarms of ants were seen dotted away from parent nests, forming new and transient nests. This phenomenon was reported by Adams (1993), and Welch (1978) who showed that only three nests occupied the same position within a woodland in Kent during ten years. In the area under the pylons, nests should show a longer site fidelity because there is a climax community of herbs and shrubs that have become established during the 60 years that coppicing has been carried out in the area. In Pound Wood all the nests that were previously abandoned were re-colonised during spring, which may have been because ants return to the parent nest during the winter when resources such as invertebrates are sparse.
Further studies on the factors affecting the distribution of Formica rufa could include carrying out similar studies on the colonies of wood ants inhabiting Little Haven Nature Reserve adjacent to Pound Wood, enabling the conclusions drawn from this study to be explored. Pound Wood supports a flourishing population of wood ants, which will continue to grow as long as the wood does not fall into neglect. It would be interesting to follow the nest movements within the wood over a period of time, in a similar study to the one carried out by Welch (1978). The movement of nests would give information about the preferred habitat and microclimate of the ant, and also test the hypothesis that movement by nests reflects the coppice cycle. Formica rufa has received much attention as a ‘keystone species’ because of the changes in the guild structure of insects on trees that it forages (Fowler and Macgarvin 1985). It would be very interesting to conduct further studies on the foraging patterns and diet of the ants in Pound Wood, and compare the habits of Formica rufa in Pound Wood to other woodland types.
It can be concluded that the red wood ant was shown to require well drained soils on which to construct nests. In addition, Oak trees as a source of invertebrate food, were shown to be important in the success of nests. However, four factors were found to directly influence the distribution of Formica rufa within Pound Wood:
The availability of sunlight
The age of coppice understorey
The position of competing nests
Tree stumps or other wooden structures on which nests were built